Biomédica 2017;37(Supl.2):77-82
doi: https://doi.org/10.7705/biomedica.v34i2.3444
ARTÍCULO ORIGINAL
1 Carrera de Medicina, Centro Universitario del Sur, Universidad de Guadalajara, Ciudad Guzmán, Jalisco, México
2 Facultad de Medicina, Universidad Autónoma de Chihuahua, Chihuahua, Chihuahua, México
3 Laboratorio de Entomología Médica, Centro Universitario del Sur, Universidad de Guadalajara, Ciudad Guzmán, Jalisco, México
Contribución de los autores:
José Alejandro Martínez-Ibarra: supervisión del equipo de trabajo, diseño del estudio y análisis de resultados
ángel Licón y Mixtli Astorga: diseño del estudio y análisis de resultados
Todos los autores participaron en las recolecciones en campo, el trabajo de laboratorio y la escritura del manuscrito.
Recibido: 23/08/16; aceptado: 23/06/17
Introducción. Los híbridos de especies y subespecies de triatominos mexicanos exhiben características biológicas y de comportamiento sobresalientes comparadas con las de sus parentales, específicamente su vigor híbrido, lo cual puede incrementar el riesgo de transmisión de Trypanosoma cruzi a sus huéspedes humanos y animales.
Objetivo. Describir y comparar los valores de los parámetros biológicos relacionados con el ciclo de vida de Triatoma recurva, de Meccus phyllosomus phyllosomus y de sus híbridos de laboratorio, para estimar su riesgo como transmisores de T. cruzi a las poblaciones humanas.
Materiales y métodos. Se entrecruzaron ejemplares de T. recurva y de M. p. phyllosomus y se obtuvo una cohorte híbrida. Las tres cohortes se mantuvieron bajo condiciones similares de laboratorio. En las tres se compararon los datos de sus ciclos de vida, el número de alimentaciones necesarias para mudar, el porcentaje de mortalidad, el de eclosión de huevos y la fecundidad de las hembras.
Resultados. La cohorte híbrida tuvo el menor tiempo promedio entre el primer instar o estadio y el adulto (136,6 ± 17,5 días). El número de alimentaciones para cambiar de instar (13,4 ± 1,7; 15,6 ± 5,2) fue mayor en la cohorte híbrida y en la de T. recurva. La cohorte híbrida registró la menor mortalidad (29 %) y el mayor porcentaje (70 %) de hembras al final del ciclo biológico. La fecundidad por hembra por día (rango: 1,1 ± 0,6 -1,6 ± 0,7) fue similar entre las poblaciones. El porcentaje de eclosión de huevos fue superior a 70 % en la cohorte híbrida y en la de M. p. phyllosomus.
Conclusión. La cohorte híbrida presentó algunas características biológicas que implicarían un mayor poder transmisor de T. cruzi que el de sus grupos parentales.
Palabras clave: Triatominae; Trypanosoma cruzi; enfermedad de Chagas; estadios del ciclo de vida; México
doi: https://doi.org/10.7705/biomedica.v34i2.3444
Biological parameters of Meccus phyllosomus phyllosomus(Burmeister), 1835, Triatoma recurva (Stål), 1868 (Hemiptera, Reduviidae) and their laboratory hybrids
Introduction: Hybrids from species and subspecies of Mexican triatomines have shown outstanding biological and behavioral characteristics as compared with their parental groups. This could lead to an increasing risk of transmission of Trypanosoma cruzi to human and animal reservoir hosts.
Objective: To describe and compare biological parameters related to the life cycle of Triatoma recurva, Meccus phyllosomus phyllosomus and their laboratory hybrids and estimate their risk as transmitters of T. cruzi to humans.
Materials and methods: We kept a cohort of each one of the three groups (T. recurva, M. p. phyllosomus and their hybrids) under similar laboratory conditions and then compared them to record the data related to their life cycles, number of blood meals to molt, mortality rates, egg eclosion rates and female fecundity.
Results: The hybrid cohort showed the lowest mean time from first-instar nymph to adult (136.6 ± 17.5 days). The number of blood meals from molt to next instar (13.4 ± 1.7; 15.6 ± 5.2) was higher in the hybrid and T. recurva cohorts. The hybrid cohort showed the lowest mortality rate (29.5%) and the highest rate (70%) of adult females at the end of the biological cycle. Fecundity per day per female (range: 1.1 ± 0.6– 1.6 ± 0.7 eggs) was similar in the three cohorts. Egg hatching rate was higher than 70% in both hybrid and M. p. phyllosomus cohorts.
Conclusion: The hybrid cohort showed some outstanding biological characteristics, which would entail a higher risk of transmission of T. cruzi to hosts than their parentals.
Key words: Triatominae; Trypanosoma cruzi; Chagas disease; life cycle stages; Mexico.
doi: https://doi.org/10.7705/biomedica.v34i2.3444
En México existen, al menos, 32 especies de triatominos. De estas, nueve se consideran las de mayor importancia epidemiológica (1),incluidas cinco de las seis que integran el revalidado género Meccus (2-4), pues se estima que son responsables de 74 % de la transmisión vectorial de Trypanosoma cruzi a los seres humanos en México (5).
Dado que existe una controversia sobre el nivel taxonómico (especies o subspecies) de los seis grupos de Meccus, se sugiere consultar una publicación reciente (6) en la cual se esclarece un tanto el tema. En este contexto, en el presente trabajo uno de los grupos estudiados fue Meccus phyllosomus phyllosomus, grupo que se encuentra distribuido en el sur de México, donde se le considera de importancia epidemiológica.
Asimismo, entre las nueve especies mexicanas de más importancia está Triatoma recurva, considerada un vector importante en las zonas norte y noroeste de México (1). Dado que algunos análisis moleculares (7,8) han establecido que T. recurva es filogenéticamente cercana a las subspecies de Meccus, en un estudio se entrecruzaron (incluidos cruces inversos) estas seis subspecies con T. recurva, con un único cruce exitoso: el de ♀ T. recurva y ♂ M. p. phyllosomus (9).
En dos estudios recientes (10,11) sobre los parámetros biológicos de los híbridos de grupos filogenéticamente cercanos, se ha demostrado el vigor Las parejas entrecruzadas se mantuvieron bajo una híbrido de los descendientes, con el consecuente aumento del riesgo de transmisión de T. cruzi a las poblaciones humanas. Por ello, el objetivo de este estudio fue conocer los parámetros biológicos de los parentales de T. recurva, M. p. phyllosomus y de sus descendientes.
Materiales y métodos
Material biológico
Para este estudio se utilizaron dos colonias de triatominos establecidas desde 2014; se recolectó un mínimo de 30 individuos procedentes de refugios de animales domésticos y de granja en el peridomicilio en dos localidades ubicadas en diferentes regiones de México. Triatoma recurva se recolectó en Satevó, Chihuahua (26° 58’ N, 107° 42 ’O), en tanto que los individuos de M. p. phyllosomus procedían de Tehuantepec, Oaxaca (16° 28’ N, 95° 19’ O).
Los ejemplares recolectados en las áreas descritas se identificaron mediante las claves taxonómicas de uso más común (12), teniendo en cuenta la revalidación del género Meccus (2).
Entrecruzamientos
Se hicieron cuatro sets de cruces. En el set A se entrecruzaron individualmente 10 hembras de T. recurva con 10 machos de M. p. phyllosomus. En el set B se hizo el cruce inverso: 10 machos de T. recurva con 10 hembras de M. p. phyllosomus, aunque en un estudio anterior se detectó su inviabilidad. Los sets C y D incluyeron los cruces parentales de 10 hembras y 10 machos de T. recurva y de 10 hembras y 10 machos de M. p. phyllosomus, respectivamente.
Parámetros biológicos
Las parejas entrecruzadas se mantuvieron bajo una temperatura de 25 ± 1o C, una humedad relativa de 70 ± 5 % y un fotoperiodo de 12 horas. A todos los ejemplares se les alimentó quincenalmente sobre conejos Nueva Zelanda inmovilizados, previamente anestesiados con 0,25 ml/kg de ketamina, en cumplimiento de la Norma Oficial Mexicana (13) y los lineamientos del Comité de ética del Centro Universitario del Sur de la Universidad de Guadalajara.
Se seleccionaron conejos como fuente de alimentación dado que en estudios previos (14,15) con T. recurva y M. p. phyllosomus, se ha concluido que son una buena fuente de alimentación sanguínea, ya que favorecen ciclos cortos en los triatominos, así como otros parámetros biológicos.
Se recolectaron los huevos de cada colonia hasta obtener 200 de cada una. Después de la eclosión, se conformaron grupos de 10 ninfas de cada población, nueve de las cuales pudieron marcarse distintiva e individualmente en el pronoto y el conexivo con marcador rojo indeleble como sigue: cuadrante superior izquierdo, cuadrante superior derecho, cuadrante inferior izquierdo, cuadrante inferior derecho, cuadrantes superiores, cuadrantes inferiores, cuadrantes laterales izquierdos, cuadrantes laterales derechos o todo el insecto. La ninfa restante se pintó totalmente con marcador azul indeleble. Las ninfas marcadas se colocaron dentro de vasos de plástico de 5,5 cm de diámetro x 10,5 cm de alto, a los cuales se les colocó un papel doblado en forma de acordeón para soporte.
Se observó a las ninfas al final de la alimentación con el fin de registrar individualmente si había ocurrido la ingestión de sangre. Los insectos se llevaron a una incubadora a 25 ± 1 °C de temperatura y 70 ± 5 % de humedad con un fotoperiodo de 12 horas; diariamente se revisó a cada individuo, con el fin de registrar las mudas y las muertes. Al finalizar el ciclo biológico, se registró la proporción de hembras y machos de cada cohorte.
Entre los ejemplares que completaron el desarrollo hasta su fase adulta, se escogieron diez parejas de cada cohorte y cada par se colocó en un recipiente plástico similar al usado con las ninfas y se las crio bajo las condiciones previamente descritas para registrar los datos de oviposición. Los huevos se contaron a lo largo de 90 días y se colocaron en recipientes separados por día de oviposición y por cohorte hasta su eclosión.
Ciclo de vida. Para calcular la duración promedio del ciclo biológico, se estableció la media de duración en días para cada estadio de desarrollo de la siguiente el estadio de ninfa de cada uno de los individuos (incluidos los de menor y de mayor duración) dividida por el número de individuos en el estadio. El número de alimentaciones hasta la muda se obtuvo dividiendo el número total de alimentaciones que cada individuo requirió para llegar a adulto por el número de ejemplares estudiados por cohorte.
Análisis estadístico
En la comparación de la cantidad de huevos por hembra, el ciclo de vida y el número de alimentaciones, primero se hizo un análisis multivariado mediante el método de Holm-Sidak, y se encontró que las distribuciones eran normales, por lo que se procedió a hacer el análisis de varianza (ANOVA).
En la comparación de los porcentajes de mortalidad entre los grupos de estudio, se utilizó la prueba de ji al cuadrado. Las diferencias se consideraron significativas cuando p resultó menor de 0,05. Los análisis se hicieron con el paquete estadístico Sigma Stat, versión 3.1 para Windows (Systat Software Inc., San José, California, Estados Unidos).
Resultados
Del set de cruces B (10 machos de T. recurva con 10 hembras de M. p. phyllosomus) no se obtuvo ni un solo huevo fértil, por lo cual no fue posible obtener información sobre sus parámetros biológicos. De los cruces de los tres sets restantes, sí se obtuvo descendencia.
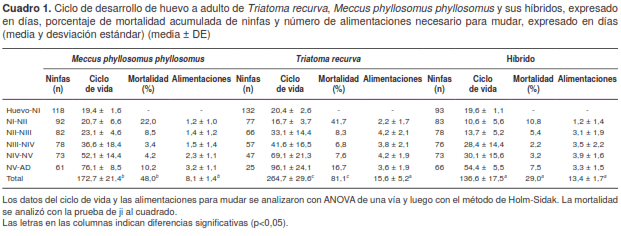
El ciclo de vida de las tres cohortes en estudio fue significativamente diferente (t=6,06, t=5,55, gl=2, p<0,05); la cohorte híbrida y la parental de T. recurva tuvieron el menor tiempo y el más largo, respectivamente (cuadro 1). El número medio de alimentaciones requerido para mudar del primer instar o estadio a adulto varió entre 8 y 21 veces, siendo significativamente menor únicamente en la cohorte de M. p. phyllosomus (t=7,45, t=5,99, gl=2, p<0,05) (cuadro 1).
El porcentaje de mortalidad varió de casi la tercera parte de los ejemplares híbridos a un poco más de 80 % en T. recurva, registrándose diferencias significativas (c2=13,57, c2=15,98, c2 =17,59, gl=1, p<0,05) entre las tres cohortes estudiadas (cuadro 1).
El porcentaje de hembras fue significativamente mayor (c2=15,24, c2=16,31, gl=1, p<0,05) en la cohorte híbrida que en las dos parentales, en las cuales no difirió (c2 =0,04 gl=1, p<0,05). Un fenómeno similar ocurrió con los machos (c2=14,15, c2=12,95, gl=1, p<0,05) (cuadro 2).
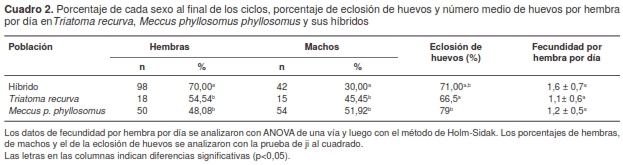
El porcentaje de eclosión de huevos varió significativamente (c2=9,46, c2=1,gl=1,p<0,05) entre las cohortes de T. recurva y las de M. p. phyllosomus, en las cuales no se registró una diferencia significativa con respecto a la cohorte híbrida (cuadro 2).
La fecundidad por hembra por día, por el contrario, fue similar (t=1,81, t=1,59, gl=1, p>0,05) en las tres cohortes estudiadas (cuadro 2).
Discusión
La duración del ciclo de vida en las dos cohortes parentales fue similar a lo reportado previamente para dichos grupos (14,15). Sin embargo, el ciclo de vida de los híbridos fue de menor duración que el de ambos parentales, incluido el ciclo más corto de M. p. phyllosomus. Un fenómeno similar se reportó en un estudio en el cual se compararon los híbridos de tres subspecies de M. phyllosomus (pallidipennis, mazzottii y phyllosomus) con esta última subspecie (11). La disminución del ciclo biológico de una especie resulta en su rápido incremento, pues cuantos más adultos haya, mayor es la abundancia de la población. Ese rápido incremento de la población de triatominos en un área podría generar, a su vez, un incremento en el riesgo de transmisión de T. cruzi a las poblaciones humanas.
Entre 65 y 75 % de los ejemplares de la cohorte de M. p. phyllosomus requirió, en promedio, una toma y media de sangre para mudar al siguiente estadio, en tanto que las cohortes de T. recurva y la híbrida requirieron de tres tomas. La necesidad de más alimentaciones conlleva, necesariamente, una mayor interacción entre vector y huésped, con el consecuente incremento del riesgo de transmisión de T. cruzi, como ocurrió en el caso de la cohorte de T. recurva y en la híbrida (16).
Los porcentajes de mortalidad de las cohortes de T. recurva y de M. p. phyllosomus fueron similares a los de estudios previos (11,14,15), lo cual contribuye a explicar la significativa presencia de M. p. phyllosomus en su área de distribución y la poca abundancia de T. recurva en gran parte de la suya (17,18). Por su parte, la cohorte híbrida mostró un porcentaje de mortalidad inferior a 30 %, lo cual podría resultar en una gran abundancia de híbridos de los dos grupos bajo estudio, con el consiguiente riesgo de transmisión de T. cruzi debido, además, a la capacidad vectorial de dichos híbridos (Memorias de la Semana Nacional de Investigación Científica-CUSur, 2015).
La proporción entre hembras y machos fue de alrededor de 1:1 en las cohortes parentales estudiadas. Aparentemente, dichas cohortes tendrían el mismo potencial de incrementar su abundancia bajo condiciones favorables. Por otra parte, en la cohorte híbrida la proporción de hembras y machos fue de 2:1, lo cual sugeriría que su potencial para aumentar sus poblaciones es mayor, dado que cuanto mayor sea la cantidad de hembras que ponen huevos, mayor será la cantidad de individuos en la población, y ello podría resultar en un incremento del riesgo de transmisión vectorial de T. cruzi a los huéspedes en sus áreas de distribución.
El porcentaje de eclosión de los huevos fue mayor en la cohorte de M. p. phyllosomus que en las otras dos, sin embargo, solo difirió de manera importante en la cohorte de T. recurva. La cohorte híbrida mostró un porcentaje de eclosión significativo y similar a su parental M. p. phyllosomus, así como al de varias especies y subespecies consideradas como importantes vectores de T. cruzi, a saber: T. patagonica, T. ryckmani, M. p. picturatus y M. p. mazzottii (6,11,19,20).
Si bien la fecundidad por hembra por día no difirió de manera importante entre las tres cohortes bajo estudio, sí fue ligeramente mayor en la cohorte híbrida, confiriéndole una ventaja con respecto a las cohortes parentales. Estos resultados fueron similares a los obtenidos en la comparación hecha en estudios separados entre M. p. pallidipennis, M. p. longippennis y M. p. picturatus y sus híbridos, y entre M. p. pallidipennis, M. p. mazzottii y M. p. phyllosomus, y sus respectivos híbridos (6,11).
Los resultados más favorables de la cohorte híbrida en la mitad de los parámetros biológicos estudiados indican que esta presenta un fenómeno de heterosis o “vigor híbrido” (21), el cual podría incrementar el riesgo epidemiológico y facilitar la aparición de tripanosomiasis, como se ha registrado en varios países suramericanos con diversas especies (22,23).
La información obtenida en este y otros estudios anteriores (6,10,11) sobre híbridos mexicanos, resalta la necesidad de estudiar la presencia de híbridos en condiciones de campo y conocer su potencial como transmisores de T. cruzi.
Conflicto de intereses
Los autores del presente escrito declaran que no existe conflicto de intereses.
Financiación
Este proyecto fue financiado por la Universidad de Guadalajara, proyecto UDG-CUSUR-SA/CIP/009/2015/. Asimismo, se recibieron fondos de la Secretaría de Educación Pública a través del proyecto 230128 UDG-CA-621: “Epidemiología de la tripanosomiasis americana en la región occidente de México”.
Correspondencia:
José Alejandro Martínez-Ibarra, Laboratorio de Entomología Médica, Centro Universitario del Sur, Universidad de Guadalajara, Avenida Enrique Arreola Silva 884, 49000 Ciudad Guzmán, Jalisco, México
Teléfono y fax: (52) (341) 575 2222
1. Ramsey JM, Townsend-Peterson A, Carmona-Castro ó, Moo-Llanes DA, Nakazawa Y, Butrick M, et al. Atlas of Mexican Triatominae (Reduviidae: Triatominae) and vector transmission of Chagas disease. Mem Inst Oswaldo Cruz. 2015;110:339-52. https://doi.org/10.1590/0074-02760140404
2. Carcavallo R, Jurberg J, Lent H, Noireau F, Galvão C. Phylogeny of the Triatominae (Hemiptera: Reduviidae): Proposal for taxonomic arrangements. Entomol Vect. 2000;7(Suppl.1):1-99.
3. Bargues MD, Klisiowicz DR, González-Candelas F, Ramsey JM, Monroy C, Ponce C, et al. Phylogeography and genetic variation of Triatoma dimidiata, the main Chagas disease vector in Central America and its position within the genus Triatoma. PLoS Negl Trop Dis. 2008;2:e233. https://doi.org/10.1371/journal.pntd.0000233
4. Bargues MD, Zuriaga Má, Mas-Coma S. Nuclear rDNA pseudogenes in Chagas disease vectors; evolutionary implications of a new 5.8S+ITS-2 paralogous sequence marker in triatomines of North, Central and northern South America. Infect Genet Evol. 2014;21:134-56. https://doi.org/10.1016/j.meegid.2013.10.028
5. Ibarra-Cerdeña CN, Sánchez-Cordero V, Townsend Peterson A, Ramsey JM. Ecology of North American Triatominae. Acta Trop. 2009;110:178-86. https://doi.org/10.1016/j.actatropica.2008.11.012
6. Martínez-Ibarra JA, Nogueda-Torres B, Cárdenas De la Cruz Má, Villagrán ME, de Diego-Cabrera JA, Bustos-Saldaña R. Biological parameters of interbreeding subspecies of Meccus phyllosomus (Hemiptera: Reduviidae: Triatominae) in western México. Bull Entomol Res. 2015;105:763-70. https://doi.org/10.1017/S0007485315000759
7. Pfeiler E, Bitler BG, Ramsey JM, Palacios-Cardiel C, Markow TA. Genetic variation, population structure, and phylogenetic relationships of Triatoma rubida and T. recurva (Hemiptera: Reduviidae: Triatominae) from the Sonoran desert, insect vec tors of the Chagas’ disease parasite Trypanosoma cruzi. Mol Phylogenet Evol. 2006;41:209-21. https://doi.org/0.1016/j.ympev.2006.07.001
8. Espinoza-Gutiérrez B, Martínez-Ibarra JA, Villalobos G, De la Torre P, Laclette JP, Martínez F. Genetic variation of North American Triatomines (Insecta: Hemiptera: Reduviidae) initial divergence between species and populations of Chagas disease vector. Am J Trop Med Hyg. 2013;88:275-84. https://doi.org/10.4269/ajtmh.2012.12-0105
9. Martínez-Ibarra JA, Nogueda-Torres B, Licón-Trillo á, Alejandre-Aguilar R, Salazar-Schettino PM, Vences Blanco MO. Biological aspects of crosses between Triatoma recurva and other members of the Phyllosoma complex. J Vect Ecol. 2015;40:117-22. https:/doi.org/10.1111/jvec.12133
10. Martínez-Ibarra JA, Nogueda-Torres B, García-Lino JC, Arroyo-Reyes D, Salazar-Montaño LF, HernándezNavarro Já, et al. Importance of hybrids of Meccus phyllosomus mazzottii, M. p. pallidipennis and M. p. phyllosomus in transmission of Trypanosoma cruzi in México. Jpn J Infect Dis. 2016;69:202-6. https://doi.org/10.7883/yoken.JJID.2015.111
11. Martínez-Ibarra JA, Nogueda-Torres B, Salazar-Montaño LF, García-Lino JC, Arroyo-Reyes D, Hernández-Navarro Já. Comparison of biological fitness in crosses between subspecies of Meccus phyllosomus (Hemiptera: Reduviidae: Triatominae) in southern México. Insect Sci. 24:114-21. https://doi.org/10.1111.1744-7917.12246
12. Lent H, Wygodzinsky P. Revision of the triatominae (Hemiptera: Reduviidae) and their significance as vectors of Chagas disease. Bull Am Mus Nat Hist.1979;163:123-520.
13. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. Norma Oficial Mexicana NOM-062ZOO-1999. Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio – 1999. Fecha de consulta: 9 de julio de 2016. Disponible en: http://www.fmvz.unam.mx/fmvz/principal/archivos/062ZOO.pdf.
14. Martínez-Ibarra JA, Alejandre-Aguilar R, Torres-Morales A, Trujillo-García JC, Nogueda-Torres B, Trujillo-Contreras F. Biology of three species of the Meccus phyllosomus complex (Hemiptera: ReduviidaeTriatominae) fed on blood of hens and rabbits. Mem Inst Oswaldo Cruz. 2006;101:787-94. https://doi.org/10.1590/S0074-02762006000700014
15. Martínez-Ibarra JA, Paredes-González E, Licón-Trillo á, Montañéz-Valdéz OD, Rocha-Chávez G, NoguedaTorres B. The biology of three Mexican-American species of Triatominae (Hemiptera: Reduviidae Triatominae): Triatoma recurva, Triatoma protracta and Triatoma rubida. Mem Inst Oswaldo Cruz. 2012;107:659-63. https://doi.org/10.1590/S0074-02762012000500013
16. Rocha DS, Jurberg J, Carcavallo RU, Presgrave OA, Cunha V, Galvão C. Influence of temperature and humidity on the nymphal de velopment of Rhodnius robustus. Rev Saúde Pública. 2001;35:400-6. https://doi.org/10.1590/S0034-89102001000400011
17. Licón-Trillo A, Balsimelli-De La Peña K, Acosta-Legarda M, Leal-Solís I, Nogueda-Torres B, Martínez-Ibarra JA. Infección natural por Trypanosoma cruzi en triatominos del Centro y Norte de México. Bol Mal Salud Amb. 2010;50:311-3.
18. Martínez-Ibarra JA, Montañéz-Valdéz OD, Rocha-Chávez G, Tapia-González JM, Nogueda-Torres B. Presencia de Meccus longipennis y Triatoma recurva en el estado de Durango, México. Bol Mal Salud Amb. 2012;52:129-33.
19. Zeledón R, Cordero M, Marroquín R, Seixas-Lorosa E. Life cycle of Triatoma ryckmani (Hemiptera: Reduviidae) in the laboratory, feeding patterns in nature and experimental infection with Trypanosoma cruzi. Mem Inst Oswaldo
Cruz. 2010;105:99-102. https://doi.org/10.1590/S007402762010000100015
20. Nattero J, Rodríguez CS, Crocco L. Effects of blood source on food resource use and reproduction in Triatoma patagonica Del Ponte (Hemiptera, Reduviidae). J Vect Ecol. 2013;38:127-33. https://doi.org/10.1111/j.1948-7134. 2013.12018.x
21. Kumar-Baranwal V, Mikkilineni V, Usha Barwale-Zehr U, Tyagi AK, Kapoor S. Heterosis: Emerging ideas about hybrid vigour. J Exp Bot. 2012;63:6309-14. https://doi.org/10.1093/jxb/ers291
22. Mas-Coma S, Bargues MD. Populations, hybrids and the systematic concepts of species and subspecies in Chagas disease triatomine vectors inferred from nuclear ribosomal and mitochondrial DNA. Acta Trop. 2009;110:112-36. https://doi.org/10.1016/j.actatropica.2008.10.013
23. Almeida CE, Oliveira HJ, Galvão C. Dispersion capacity of Triatoma sherlocki, Triatoma juazeirensis and laboratorybred hybrids. Acta Trop. 2012;122:71-9. https://doi.org/10.1016/j.actatropica.2011.12.001