Biomédica 2017;37(Supl.2):59-66
doi: https://doi.org/10.7705/biomedica.v34i2.3471
ARTÍCULO ORIGINAL
1 Grupo de Epidemiología y Salud Poblacional, Universidad del Valle, Cali, Colombia
2 Secretaría de Salud Pública Municipal, Cali, Colombia
3 Grupo de Investigación Biomédica, Universidad Icesi, Cali, Colombia
4 Grupo de Saneamiento Ambiental, Instituto CINARA, Universidad del Valle, Cali, Colombia
Contribución de los autores:
Daniel Elías Cuartas, Miguel Peña y Fabián Méndez: diseño del estudio, análisis e interpretación de los datos
Genny Martínez y Yoseth Ariza-Araújo: diseño del estudio y recolección de información
Diana María Caicedo, Yoseth Ariza-Araújo y Jhonny Garcés: análisis e interpretación de los datos
Todos los autores participaron en la escritura del manuscrito.
Recibido: 08/08/16; aceptado: 08/06/17
Introducción. La distribución espacial de Aedes aegypti es heterogénea, y la interacción entre criaderos positivos y potenciales en el intradomicilio y el extradomicilio es uno de los aspectos más difíciles de caracterizar en los programas de control vectorial.
Objetivo. Describir la relación espacial entre los criaderos potenciales y positivos de A. aegypti en el intradomicilio y en el extradomicilio en un sector de Cali, Colombia.
Materiales y métodos. Se hizo una encuesta entomológica con el objetivo de recolectar datos de los criaderos en el intradomicilio y el extradomicilio. El análisis exploratorio de los datos espaciales incluyó la localización, la tendencia espacial, la autocorrelación espacial local, la continuidad espacial y la correlación espacial de los criaderos positivos y potenciales según el hábitat.
Resultados. Se determinaron las tendencias espaciales. Mediante el análisis de autocorrelación espacial local se ubicaron los conglomerados de criaderos potenciales y positivos del extradomicilio. Se encontró una correlación positiva entre los criaderos potenciales y los positivos y una correlación negativa entre el intradomicilio y el extradomicilio.
Conclusiones. La relación espacial entre criaderos positivos y potenciales de A. aegypti en el intradomicilio y el extradomicilio es dinámica y muy sensible a las características de cada territorio, por lo que establecer su distribución en el espacio contribuye a la priorización de recursos y acciones en los programas de control vectorial.
Palabras clave: Aedes aegypti, dengue, Zika, chikungunya, análisis espacial.
doi: https://doi.org/10.7705/biomedica.v34i2.3471
Spatial distribution of potential and positive Aedes aegypti breeding sites
Introduction: The spatial distribution of Aedes aegypti is heterogeneous, and the interaction between positive and potential breeding sites located both inside and outside homes is one of the most difficult aspects to characterize in vector control programs.
Objective: To describe the spatial relationship between potential and positive breeding sites of A. aegypti inside and outside homes in Cali, Colombia.
Materials and methods: We conducted an entomological survey to collect data from both indoor and outdoor breeding sites. The exploratory analysis of spatial data included location, spatial trends, local spatial autocorrelation, spatial continuity and spatial correlation of positive and potential breeding sites according to habitat.
Results: Spatial trends were identified, as well as clusters of potential and positive breeding sites outdoors using local spatial autocorrelation analysis. A positive correlation was found between potential and positive breeding sites, and a negative correlation existed between indoor and outdoor sites.
Conclusions: The spatial relationship between positive and potential A. aegypti breeding sites both indoors and outdoors is dynamic and highly sensitive to the characteristics of each territory. Knowing how positive and potential breeding sites are distributed contributes to the prioritization of resources and actions in vector control programs.
Key words: Aedes aegypti, dengue, Zika, Chikungunya, spatial analysis.
doi: https://doi.org/10.7705/biomedica.v34i2.3471
El dengue, el Zika y el chikungunya son enfermedades infecciosas causadas por arbovirus (1-3) y transmitidas por dípteros del género Aedes (4). La complejidad de la transmisión de estas enfermedades (5-7) supera los aspectos biológicos de la interacción entre mosquitos y seres humanos, e involucra factores determinantes, como el ambiente urbano, los comportamientos individuales y grupales de los habitantes de las ciudades y las formas particulares de organización de los servicios de control y prevención.
Según la Organización Panamericana de la Salud (8), el control entomológico de las poblaciones del mosquito es de primordial importancia e involucra la fumigación periódica y la eliminación de criaderos positivos y potenciales en el marco de los programas de control y prevención. Los criaderos positivos son aquellos que tienen pupas o larvas y, los criaderos potenciales, aquellos que, a pesar de no contener pupas o larvas, tienen la capacidad de retener agua y, por su localización en el interior de la vivienda o en sus alrededores, son de fácil acceso para la oviposición de las hembras de A. aegypti.
Cali es la tercera ciudad más importante de Colombia y un lugar de transmisión hiperendémica del dengue, a lo cual se añade ahora el surgimiento del Zika y el chikungunya. Los sumideros (alcantarillas) de la ciudad son el principal criadero positivo y potencial de A. aegypti. Estas estructuras urbanas de manejo y control de las aguas lluvia, han sido colonizadas por A. aegypti y pueden aportar entre 40 y 495 pupas por manzana, lo que, sumado a los criaderos situados en el intradomicilio, puede generar una población de mosquitos adultos que varía entre 45 y 555 individuos por manzana (9).
La combinación de factores ambientales y sociales hace que la distribución de la población de mosquitos no sea homogénea en tiempo y espacio; además, la interacción entre criaderos positivos y potenciales en el interior y exterior de las viviendas en un territorio específico constituye uno de los de aspectos más difíciles de caracterizar en el manejo y control de criaderos del vector.
En este sentido, el presente artículo se propuso describir la relación espacial entre criaderos positivos y potenciales en el intradomicilio y en el extradomicilio a partir de un análisis exploratorio de datos espaciales y del análisis de correlación.
Materiales y métodos
Área de estudio
Cali tiene una población cercana a los 2,5 millones de habitantes y está a una altitud entre los 950 y 1.070 msnm en un valle interandino, con un ecosistema de bosque seco tropical, una temperatura promedio de 24,6 °C y pluviosidad promedio de 1.588 mm (10), condiciones ideales para A. aegypti.
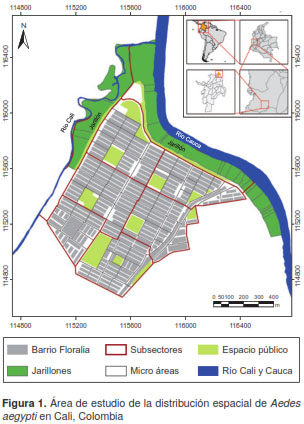
Una de las áreas con mayor presencia de este vector se encuentra en el nororiente de la ciudad, en una zona con características particulares, pues se ubica en la desembocadura del río Cali en el río Cauca y cuenta con dos tipos de desarrollo urbano: el planificado, por un lado, y, por el otro, los asentamientos informales sobre farillones o ‘jarillones’ (diques o terraplenes que protegen la ciudad de inundaciones) de los ríos Cali y Cauca.
Para la recolección detallada de información, el área de estudio se dividió en dos sectores según sus características y la morfología urbana. El primer sector correspondió al área periurbana de conformación subnormal situada sobre los farillones de los ríos Cauca y Cali, los cuales se emplearon como límites para subdividir el área en dos subsectores y estos, a su vez, en 24 microáreas. El segundo sector correspondió a un área residencial planificada, que se subdividió en seis subsectores y 77 microáreas. Las microáreas de los dos sectores incluían entre una y seis manzanas (figura 1).
Recolección de la información
Para la recolección de información se definió como intradomicilio a todo el espacio de la puerta de acceso a la vivienda hacia adentro, y el extradomicilio como aquel ubicado de la puerta de acceso hacia afuera. En el caso del extradomicilio se hizo un único recorrido a lo largo de los 174 lados de las 101 microáreas establecidas. En el instrumento de observación diseñado, se registró la información relativa a la presencia de la vegetación que podía retener agua (árboles con axilas y bromelias), de sumideros, escombros, botaderos crónicos de basura, recipientes y llantas.
Para recolectar la información del intradomicilio, se desarrolló una única inspección entomológica en 846 viviendas (9,8 % de las viviendas del sector), de las cuales 87 (10,3 %) se localizaban en los farillones de los ríos Cali y Cauca. Las viviendas se seleccionaron por medio de un muestreo aleatorio sistemático a partir de un marco muestral finito conformado por 260 manzanas del área de estudio. En cada manzana se seleccionaron, al menos, tres viviendas, aunque en algunos sectores con mayor densidad de unidades residenciales se seleccionó un mayor número.
La detección de criaderos en el intradomicilio y el extradomicilio estuvo a cargo de personal capacitado en identificación morfológica de las formas del vector. Se siguió un proceso operativo estandarizado que establecía el uso de concentradores larvales en una única ocasión para depósitos pequeños, en tanto que, para recipientes grandes y sumideros, se empleó una red acuática de abertura rectangular en tres ocasiones con cinco minutos de intervalo. En la detección de las formas inmaduras se empleó la clave taxonómica de Forattini.
Análisis espacial
Mediante el sistema de información geográfica (SIG), se establecieron la localización, la distribución, la asociación y la interacción (11,12) de las cuatro variables de interés: criaderos positivos y potenciales en el intradomicilio, en el extradomicilio. Los datos de estas cuatro variables se agregaron en la unidad de análisis, es decir, la manzana.
Se calcularon los centroides (13) de cada manzana y, a partir de estos, se calculó un área de influencia (buffer) (14) de 100 m de radio alrededor de cada uno, como un valor intermedio entre el rango dado por el máximo valor de vuelo (160 m) y el valor promedio (56 m) (15). Una vez establecidas las zonas de influencia, se contaron todos los depósitos y sumideros positivos ubicados en ellas mediante una relación espacial de ‘contenencia’ (16), dato que se consideró como el correspondiente a los criaderos positivos en cada una de las manzanas (extradomicilio). Un procedimiento similar se siguió para el conteo de los depósitos y sumideros que no tenían pupas o larvas, los cuales se consideraron como criaderos potenciales del extradomicilio.
Se hizo un análisis de la tendencia espacial para describir el comportamiento de las variables en la unidad geográfica de análisis en el área de estudio (16-19). Para determinar la presencia de patrones espaciales en las variables, se hizo un análisis de autocorrelación espacial local para determinar el índice global de Morán, el cual indica la existencia de un patrón espacial a medida que se aproxima a 1, y el índice local de Morán, con el cual se examinó la presencia de conglomerados de valores extremos y se evaluó la significación estadística de la hipótesis sobre la disimilitud entre manzanas, y los valores extremos dentro de ella (18,20). Se empleó el programa GeoDa para el análisis (19) y, el ArcGis, para el mapeo.
Para describir la continuidad espacial de cada una de las variables, se emplearon técnicas geoestadísticas. Por tratarse de variables regionalizadas, se requirió un análisis univariado, uno bivariado y uno espacial para la detección y explicación de anomalías (21). Para determinar los cambios de cada una de las variables con respecto a la distancia, se empleó el semivariograma, cuyo fundamento es la primera ley de Tobler (22), la cual establece que “todo se relaciona con todo lo demás, pero las cosas cercanas están más relacionadas que las cosas distantes”. La varianza entre pares separados por intervalos de distancia es la semivarianza (Ÿ), calculada con la siguiente ecuación:
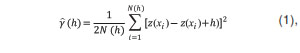
donde Ÿ(h) (h) es la semivarianza para todas las muestras localizadas en el espacio separado por el intervalo de distancia h, z(xi) y z(xi + h) son los valores de la variable observados en los puntos de muestreo, y N (h) es el número total de pares de muestra separados por un intervalo de distancia h (23).
Una vez establecido el semivariograma, se calculó la superficie continua para cada una de las variables mediante el método kriging, técnica de interpolación que proporciona el mejor estimador lineal imparcial y estima la varianza de kriging, la cual depende del modelo de variograma obtenido y de las localizaciones de los datos originales, lo que permite un análisis de la calidad de las estimaciones. Se construyeron dos modelos con cokriging, variante que permite modelar varias variables simultáneamente (24-26): uno correspondiente a la potencialidad y otro a la positividad. Se hizo validación cruzada de cada modelo y se registraron la media de los errores de estimación, el error cuadrático medio y el error cuadrático medio adimensional (27). Por último, se hizo un análisis de correlación espacial con los modelos de cada una de las variables.
Resultados
En 86 % (223) de las 260 manzanas estudiadas, se detectó como mínimo un criadero positivo y, en 98 % (256) de ellas, uno potencial en el interior de las casas. De los 210 sumideros que se evaluaron como criaderos en el extradomicilio, en 23 % (48) se encontraron larvas o pupas de A. aegypti. Al calcular los sumideros potenciales y positivos en un área de 100 m alrededor de cada manzana con base en estos datos, se encontró que en 77 % (201) de las manzanas había, por lo menos, un criadero positivo en el extradomicilio y en 96 % (251), uno potencial.
En el análisis de tendencia espacial de cada una de las variables, no se evidenció una tendencia espacial de los criaderos positivos en el intradomicilio (figura 2A), pero sí una ligera tendencia a la concentración de las manzanas con criaderos potenciales cerca del farillón del río Cauca, especialmente aquellas con valores en el tercer y el cuarto cuartiles (figura 2B).
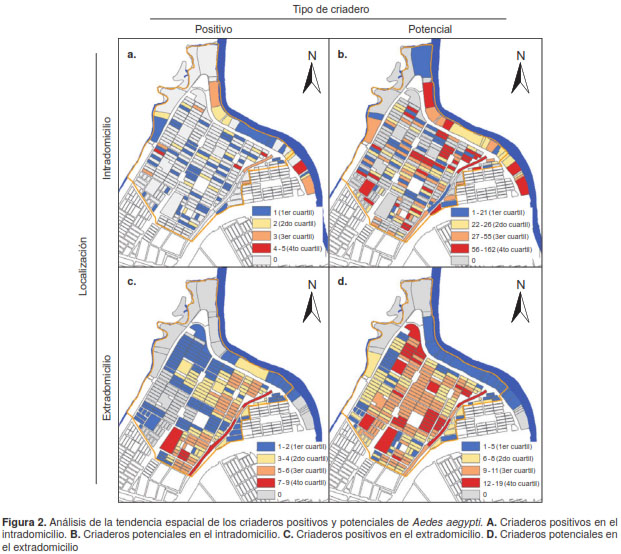
Por el contrario, en el caso de criaderos positivos en el extradomicilio, se observó una clara tendencia de ubicación hacia el occidente de la mayoría de las manzanas sin presencia de tales criaderos, o solo de algunas manzanas en el primer y segundo cuartiles, en tanto que en el oriente se concentró la mayoría de las manzanas en el tercer y cuarto cuartiles (figura 2C), en un patrón similar al de los criaderos potenciales en el extradomicilio, pero, en este caso, con tendencia a concentrarse en el centro del área de estudio (figura 2D).
Estas tendencias se comprobaron con el análisis de autocorrelación espacial local. En el interior de las viviendas, no se observó un patrón especial ni para los criaderos positivos ni para los potenciales (figura 3 A y B), pues el índice de Morán fue muy próximo a cero y, aunque algunas manzanas aparecieron como conglomerados con significación estadística, estaban aisladas y no presentaban un patrón espacial claro. En el extradomicilio, en cambio, los criaderos positivos y potenciales presentaron un patrón espacial dado por el índice de Morán (figura 3 C y D), lo cual se comprobó en el análisis de conglomerados al encontrar, en ambos casos, un conglomerado de manzanas con un gran número de criaderos rodeado de manzanas también con valores elevados, el cual resultó estadísticamente significativo.
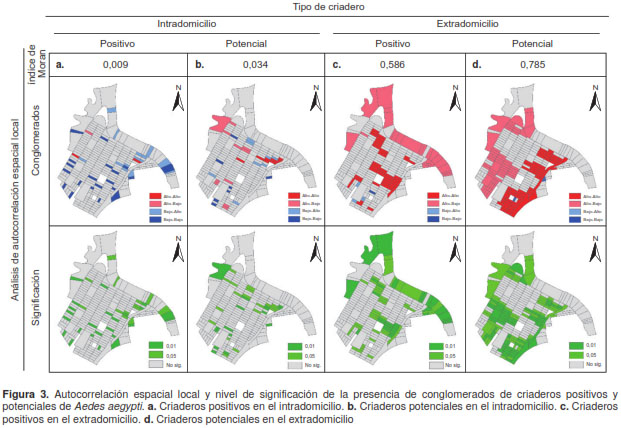
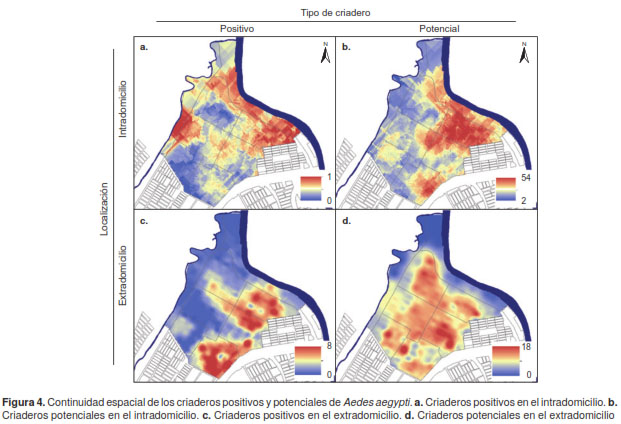
Bajo el supuesto de que la distribución de criaderos es continua en el área de estudio, debido al rango de vuelo del mosquito, se obtuvieron superficies continuas para cada uno de los tipos de criaderos. En el caso de los criaderos positivos encontrados en el intradomicilio (figura 3A), se observó que los valores altos se presentaron en el occidente y el oriente del área de estudio y que, especialmente en el occidente, coincidieron con los valores elevados de los criaderos potenciales del intradomicilio (figura 3B) y los positivos del extradomicilio (figura 3C).
En contraste, las zonas con pocos criaderos potenciales en el exterior de las viviendas se ubicaron en áreas cercanas a los farillones, o sobre ellos, en las cuales no había servicio de acueducto ni alcantarillado y, por eso, no contaban con sumideros (figura 3D); allí se registró un considerable número de criaderos positivos y potenciales en el intradomicilio. Se observó, en cambio, una clara concentración de los criaderos positivos (figura 3C) en el extradomicilio en el occidente del área de estudio, lo cual coincidió con los conglomerados detectados en el análisis de autocorrelación espacial local.
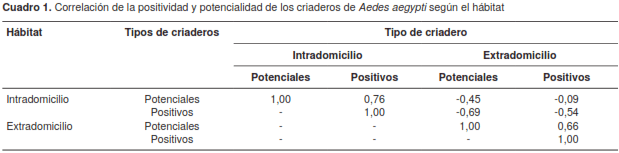
El análisis de correlación espacial de los modelos de cada una de las variables evidenció una correlación positiva de 0,76 entre los criaderos positivos y potenciales en el intradomicilio. Hubo un comportamiento similar de los criaderos positivos y potenciales del extradomicilio, los cuales se correlacionaron positivamente, con un valor de 0,66. Sin embargo, se observó una correlación negativa entre los criaderos del intradomicilio y los del extra-domicilio (cuadro 1) (figura 4).
Discusión
La relación espacial de criaderos positivos y potenciales en el interior y el exterior de las viviendas es compleja y constituye un factor clave en el ciclo vital de A. aegypti que influye en la dinámica de la transmisión de las arbovirosis para las cuales sirve de vector. En el presente artículo se exploró la relación espacial entre criaderos positivos y potenciales en el intradomicilio y el extradomicilio y se estableció su correlación espacial. Con la información recolectada en el área de estudio, se determinó la presencia de patrones espaciales para los criaderos positivos y potenciales en el extradomicilio, así como la presencia de conglomerados de manzanas con un número elevado de criaderos. Se encontró una correlación espacial positiva entre los criaderos positivos y potenciales tanto en el interior como en el exterior de las viviendas, y una correlación espacial negativa entre estos dos hábitats.
Como lo demuestran los resultados, la clasificación y el análisis de los criaderos como potenciales y positivos permitieron obtener información útil para las acciones de control y prevención del dengue, lo cual se reforzó con el análisis de la distribución espacial según las categorías de intradomicilio y extradomicilio, pues se determinaron las áreas con una mayor concentración de criaderos por manzana, lo cual contribuye a una mejor comprensión de la dinámica de las relaciones ambientales en el comportamiento de esta infección (28).
La presencia de conglomerados de manzanas con un gran número de criaderos positivos y potenciales en el extradomicilio, confirmó la importancia de los sumideros en la producción de larvas y adultos de A. aegypti (9,29,30). Además, la correlación positiva entre los dos tipos de criaderos en el extradomicilio sugiere una continua producción de mosquitos determinada por el posible relevo entre ellos, aspecto que no fue abordado en este estudio, pero que es necesario evaluar en futuras investigaciones.
Por otro lado, la correlación negativa entre los criaderos del intradomicilio y el extradomicilio se explicaría, en parte, por el efecto del control químico de los criaderos, el cual está rezagado en el tiempo (las características del extradomicilio en un tiempo t1 afectan las del intradomicilio en un tiempo t2). Asimismo, los criaderos tienen dos características: primero, pueden cambiar de positivos a potenciales y viceversa; y segundo, su capacidad de producción de pupas depende del volumen y las propiedades físicas y químicas del agua. Esta combinación de factores condiciona la productividad y su análisis puede brindar información sobre los criaderos con mayor o menor productividad (31). Los cambios en los tipos de criaderos y la productividad constituyen elementos que podrían explorarse en estudios posteriores mediante una regresión ponderada geográficamente para determinar la variabilidad espacial con la ayuda de múltiples regresiones locales basadas en los valores de las unidades espaciales (manzanas, barrios, comunas) (32,33).
Los sumideros son el principal criadero en el extradomicilio en Cali, lo cual contrasta con lo que sucede en otras ciudades, como Girardot, donde los principales criaderos son los tanques bajos y las albercas (34). Por ello, es necesario adelantar acciones intersectoriales para la implementación de sistemas innovadores de drenaje en las ciudades, de manera que no se conviertan en hábitat ideal para el mosquito (8). Asimismo, el poder de los análisis espaciales desarrollados en este estudio demuestra que las herramientas empleadas pueden fortalecer las capacidades locales para el control y la prevención del dengue.
Los autores declaramos que no tenemos conflictos de intereses.
El estudio fue financiado por el International Development Research Centre (IDRC) de Canadá y la Universidad del Valle.
Correspondencia:
Daniel Elías Cuartas, Calle 4B N° 36-140, edificio 118, oficina 215, Cali, Colombia Teléfono: (572) 554 2476; celular: (316) 326 8449 daniel.cuartas@correounivalle.edu.co
1. Horstick O, Runge-Ranzinger S, Nathan MB, Kroeger A. Dengue vector-control services: How do they work? A systematic literature review and country case studies. Trans R Soc Trop Med Hyg. 2010;104:379-86. https://doi.org/10.1016/j.trstmh.2009.07.027
2. Fauci AS, Morens DM. Zika virus in the Americas—yet another arbovirus threat. N Engl J Med. 2016;374:601-4. https://doi.org/10.1056/NEJMp1600297
3. Pialoux G, Gaüzère B-A, Jauréguiberry S, Strobel M. Chikungunya, an epidemic arbovirosis. Lancet Infect Dis. 2007;7:319-27. https://doi.org/10.1016/S1473-3099(07)70107-X
4. Cafferata ML, Bardach A, Rey-Ares L, Alcaraz A, Cormick G, Gibbons L, et al. Dengue epidemiology and burden of disease in Latin America and the Caribbean: A systematic review of the literature and meta-analysis. Value Health Reg Issues. 2013;2:347-56. https://doi.org/10.1016/j.vhri.2013.10.002
5. McLennan-Smith TA, Mercer GN. Complex behaviour in a dengue model with a seasonally varying vector population. Math Biosci. 2014;248:22-30. https://doi.org/10.1016/j.mbs.2013.11.003
6. Aguiar M, Kooi BW, Rocha F, Ghaffari P, Stollenwerk N. How much complexity is needed to describe the fluctuations observed in dengue hemorrhagic fever incidence data? Ecological Complexity. 2013;16:31-40. https://doi.org/10.1016/j.ecocom.2012.09.001
7. Epstein PR. Chikungunya fever resurgence and global warming. Am J Trop Med Hyg. 2007;76:403-4.
8. Organización Panamericana de la Salud, Organización Mundial de la Salud. Dengue: guías para el diagnóstico, tratamiento, prevención y control. La Paz, Bolivia: OPS/OMS; 2010.
9. González R, Gamboa F, Perafán O, Suárez MF, Lerma JM. Experiencia de un análisis entomológico de criaderos de Aedes aegypti y Culex quinquefasciatus en Cali, Colombia. Revista Colombiana de Entomología. 2007;33:9.
10. Alcaldía de Santiago de Cali. Cali en cifras 2011. Cali: Alcaldía de Santiago de Cali; 2012. p. 161.
11. Buzai G. Sistemas de información geográfica en geografía de la salud. En: Pickenhayn J, editor. Salud y enfermedad en geografía. Buenos Aires: Lugar Editores; 2008. p. 111-34.
12. Palaniyandi M. The environmental aspects of dengue and chikungunya outbreaks in India: GIS for epidemic control. International Journal of Mosquito Research. 2014;1:38-44.
13. Deakin RE, Birda SC, Grenfellb RI. The Centroid? Where would you like it to be? Cartography. 2002;31:153-67. https://doi.org/10.1080/00690805.2002.9714213
14. Vine MF, Degnan D, Hanchette C. Geographic information systems: Their use in environmental epidemiologic research. Environ Health Perspect. 1997;105:598-605.
15. Muir LE, Kay BH. Aedes aegypti survival and dispersal estimated by mark-release-recapture in northern Australia. Am J Trop Med Hyg. 1998;58:277-82. https://doi.org/10.4269/ajtmh.1998.58.277
16. Ministerio da Saúde do Brazil, Fundaçao Oswaldo Cruz. Introduçao à estadistística espacial para a saúde pública. Brasilia: Ministerio da Saúde; 2007.
17. Cuartas D, Ariza Y, Pachajoa H, Méndez F. Analysis of the spatial and temporal distribution of birth defects between 2004-2008 at a third-level hospital in Cali, Colombia. Colombia Médica. 2011;42:9-16.
18. Coro C. Análisis estadístico de datos geográficos en geomarketing: el programa GeoDa. Revista Distribución y Consumo. 2006;131:34-45.
19. AnselinL, Syabri I, Kho Y. GeoDa: An introduction to spatial data analysis. Geographical analysis. 2006;38:5-22. https://doi.org/10.1111/j.0016-7363.2005.00671.x
20. Anselin L. Local Indicators of Spatial Association-LISA. Geographical Analysis. 1995;27:93-115. https://doi.org/10.1111/j.1538-4632.1995.tb00338.x
21. Isaaks EH, Srivastava RM. An introduction to applied geostatistics. Oxford: Oxford University Press; 1989. p. 561.
22. Tobler WR. A computer movie simulating urban growth in the Detroit region. Economic Geographic. 1970;46:234-40.
23. Gallardo A, Maestre F. Métodos geoestadísticos para el análisis de datos ecológicos espacialmente explícitos. En: Maestre F, Alcántara A, Bonet A, editores. Introducción al análisis espacial de datos en ecología y ciencias ambientales: métodos y aplicaciones. Madrid: Universidad Rey Juan Carlos; 2008. p. 216-72.
24. Goovaerts P. Ordinary cokriging revisited. Mathematical Geology. 1998;30:21-42. https://doi.org/10.1023/A:1021757104135
25. Knotters M, Brus DJ, Oude Vosjaar JH. A comparison of kriging, co-kriging and kriging combined with regression for spatial interpolation of horizon depth with censored observations. Geoderma. 1995;67:227-46. https://doi.org/10.1016/0016-7061(95)00011-C
26. Childs C. Interpolation surfaces in arcgis spatial analyst. ArcUser. 2004:32-5.
27. Dávila JR, González J, Ocete R, López R. Descripción estadística de la distribución espacial de los huevos del mosquito verde Jacobiasca lybica (Bergenin & Zanon) (Homoptera: Cicadellidae) en viñedo: modelización y mapeo. Bol SanVeg Plagas. 2002;28:87-95.
28. Nyamah MA, Sulaiman S, Omar B. Categorization of potential breeding sites of dengue vectors in Johor, Malaysia. Trop Biomed. 2010;27:33-40.
29. Focks DA, Daniels E, Haile DG, Keesling JE. A simulation model of the epidemiology of urban dengue fever: Literature analysis, model development, preliminary validation, and samples of simulation results. Am J Trop Med Hyg. 1995;53:489-506. https://doi.org/10.4269/ajtmh.1995.53.489
30. Ocampo CB, Giraldo-Calderón GI, Pérez M, Morales CA. Evaluación del triflumurón y la mezcla de Bacillus thuringiensis más Bacillus sphaericus para el control de las formas inmaduras de Aedes aegypti y Culex quinquefasciatus en sumideros en Cali, Colombia. Biomédica. 2008;28:22433.https://doi.org/10.7705/biomedica.v28i2.93
31. Barrera R. Recomendaciones para la vigilancia de Aedes aegypti. Biomédica. 2016;36:454-62. https://doi.org/10.7705/biomedica.v36i3.2892
32. Fotheringham S, Charlton ME, Brunsdon C. Geographically weighted regression: A natural evolution of the expansion method for spatial data analysis. Environ Plan A.1998;30:1905-27.
33. Nakaya T, Fotheringham AS, Brunsdon C, Charlton M. Geographically weighted Poisson regression for disease association mapping. Stat Med. 2005;24:2695-717.
34. Alcalá L, Quintero J, González-Uribe C, Brochero H. Productividad de Aedes aegypti (L.) (Diptera: Culicidae) en viviendas y espacios públicos en una ciudad endémica para dengue en Colombia. Biomédica. 2015;35:258-68. http://dx.doi.org/10.7705/biomedica.v35i2.2567